How Do +TIPs Control Microtubule Dynamics Through Phase Separation?
Posted by Neelabh Datta, on 31 January 2025
What do you believe drives the precise regulation of microtubule dynamics in eukaryotic cells, allowing them to efficiently divide, move, and communicate? Well, at the heart of this process lies a diverse group of proteins known as microtubule plus-end tracking proteins (+TIPs), which assemble into highly regulated networks at the growing ends of microtubules. These proteins are integral to a vast range of cellular functions, from maintaining cell polarity to ensuring accurate chromosome segregation during mitosis. While the significance of +TIPs has been well established, recent research has begun sorting out their remarkable ability to undergo liquid-liquid phase separation (LLPS), a phenomenon that may hold the key to understanding their regulation and function at a molecular level.
+TIPs are evolutionarily conserved across species, spanning from yeast to mammals, and can be categorized into two main groups based on their ability to associate with microtubule plus-ends. The first group consists of autonomous +TIPs, such as the end-binding (EB) proteins, which directly bind to microtubules and are among the most widely distributed +TIPs. The second group comprises proteins that localize to microtubule ends through interactions with EB proteins. Known members of this category include cytoplasmic linker protein 170 (CLIP-170), adenomatous polyposis coli (APC), SLAINs, and the microtubule-actin crosslinking factor (MACF). Together, these proteins establish a dynamic and multivalent interaction network that regulates microtubule behavior and coordinates cellular processes such as intracellular trafficking and mitotic spindle positioning.
In the budding yeast Saccharomyces cerevisiae, one of the best-characterized +TIP networks is mediated by the protein Kar9, which functions analogously to APC, SLAINs, and MACF in higher eukaryotes. This network comprises Kar9, the EB homolog Bim1, and the CLIP-170 ortholog Bik1. These proteins collectively regulate nuclear positioning during mitosis and mating. Recent findings have demonstrated that Kar9, Bim1, and Bik1 can undergo phase separation in vitro, forming liquid-like condensates that appear to facilitate their biological roles. Among these, Bik1 has emerged as a particularly interesting player, as it has been identified as the principal driver of condensation, substantially lowering the concentration threshold required for droplet formation.
The presence of condensates in living cells has led researchers to hypothesize the existence of a specialized structure, referred to as the “+TIP body,” which dynamically tracks microtubule plus-ends. This assembly is thought to serve as a mechanical linker between microtubules and actin cables, assisting in force transmission via the actin-based motor Myo2 during mitosis. The formation and stability of the +TIP body appear to depend on multiple factors, including structured domains, linear motifs in disordered protein regions, oligomerization tendencies, and a high degree of multivalent interactions. Interestingly, similar molecular features have been implicated in the phase separation of +TIPs in more complex eukaryotic organisms, including the well-studied CLIP-170 and EB proteins.
Despite the progress made in characterizing +TIP condensates, many aspects of their organization and regulation remain elusive. To bridge this knowledge gap, researchers have focused on Bik1, a multidomain and oligomeric protein with remarkable structural and biophysical properties. Functionally, Bik1 plays an essential role in controlling microtubule dynamics and is involved in critical cellular processes such as nuclear fusion, chromosome segregation, and mitotic spindle positioning. It has been observed that Bik1 can phase separate in vitro even in the absence of binding partners or molecular crowding agents. This behavior is particularly pronounced under low-salt conditions, where the protein forms micrometer-sized droplets with unusually low density and high viscosity, setting it apart from other known condensates.
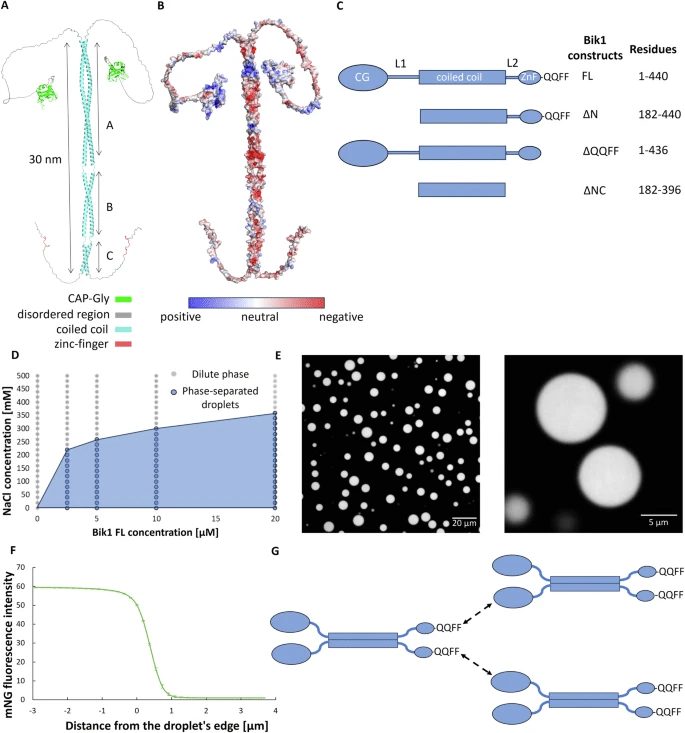
The figure shows the structural representation of the Bik1 dimer. As shown in A, AlphaFold predicts Bik1 dimer to be organized into three distinct coiled-coil segments (A, B, and C), with additional domains such as the CAP-Gly and zinc-finger motifs contributing to its functionality and flexibility, while the overall length of the protein is indicated to be approximately 30 nm. In B, the electrostatic surface charge distribution of Bik1 is mapped, where positively charged regions are marked in blue, negatively charged areas in red, and neutral zones in light gray, providing insight into potential interaction sites critical for protein-protein binding. C presents the different Bik1 truncation constructs used in the study, specifying their domain compositions and residue boundaries, including full-length (FL) Bik1, an N-terminal deletion variant (ΔN), a construct lacking the QQQF motif (ΔQQFF), and a coiled-coil-only version missing both N- and C-terminal regions (ΔNC). D illustrates the phase separation behavior of Bik1 FL in response to varying concentrations of NaCl, revealing a transition from a dilute phase to droplet formation as the protein concentration increases, while E displays confocal microscopy images showing the resulting micrometer-sized droplets formed under these conditions. Finally, F quantifies fluorescence intensity across the droplets’ edges, indicating a gradient in molecular distribution, while G proposes a model of intermolecular interactions in which Bik1 dimers are stabilized through their QQQF motifs, facilitating condensate formation and self-assembly in a dynamic cellular environment. Adapted from https://doi.org/10.1038/s41467-025-56468-8 and made available under a Creative Commons Attribution 4.0 International License.
To elucidate the structural mechanisms underlying Bik1 phase separation, an integrative approach combining computational modeling, fluorescence microscopy, small-angle X-ray scattering (SAXS), and cross-linking mass spectrometry (XL-MS) has been employed. These methods provide a holistic perspective on the protein’s conformational dynamics and intermolecular interactions in both its soluble and phase-separated states. A particularly intriguing aspect of Bik1’s self-assembly is the critical role played by its N-terminal CAP-Gly domain and C-terminal EEY/F-like motif. These regions are known to mediate interactions with α-tubulin, EB proteins, and CLIP-170 homologs. The observed phase separation of Bik1 suggests that an intramolecular interaction between its CAP-Gly domain and EEY/F-like motif may drive condensation.
SAXS data and computational analyses indicate that Bik1 adopts an elongated conformation, characterized by two kinks within its parallel coiled-coil domain. These kinks arise due to discontinuities in the heptad-repeat sequence, contributing to the protein’s overall flexibility. The CAP-Gly and C-terminal zinc-finger-containing regions are connected to the coiled-coil domain by flexible linkers, which may allow for dynamic intramolecular and intermolecular interactions. XL-MS experiments further reveal that all three coiled-coil segments of Bik1 interact with both its N- and C-terminal regions in both dilute and condensed states. Interestingly, crosslinking patterns shift significantly between high- and low-salt conditions, suggesting conformational rearrangements during phase separation.
Bik1 shares significant structural and functional similarities with its mammalian counterpart, CLIP-170. Previous studies have shown that CLIP-170 undergoes intramolecular folding due to interactions between its N-terminal CAP-Gly domain and C-terminal EEY/F motif. More recently, CLIP-170 has also been observed to form liquid-like condensates both in vitro and in living cells. It has been proposed that these condensates act as hubs for the recruitment of various proteins, including tubulin, to microtubule plus-ends. Given the functional parallels between Bik1 and CLIP-170, it is plausible that Bik1 condensation similarly facilitates microtubule regulation during yeast cell division.
To further dissect the role of specific Bik1 domains in phase separation, researchers have generated truncation mutants lacking key structural elements. Experiments reveal that while the full-length protein undergoes robust phase separation, the deletion of either the N-terminal CAP-Gly domain or the C-terminal EEY/F motif severely disrupts this ability. Moreover, a minimal truncation variant that retains oligomerization capacity but lacks the ability to form micrometer-sized droplets exhibits distinct crosslinking patterns, further supporting the importance of these domains in condensation.
Beyond characterizing individual protein components, researchers have investigated the internal organization of phase-separated Bik1 condensates. Confocal fluorescence microscopy suggests that Bik1 is homogenously distributed within droplets at a micron-scale resolution. However, SAXS measurements reveal a more complex internal architecture, with condensates exhibiting a fractal-like network organization at a length scale of 30 to 300 nm. This structural property is reminiscent of percolation networks, which emerge in polymer sol-gel transitions and have recently been proposed as a fundamental organizing principle in biomolecular condensates. The fractal structure of Bik1 condensates may explain their unique viscosity, allowing for mechanical stability while maintaining low protein concentration.
While these in vitro findings provide valuable insights into Bik1 phase separation, their implications for cellular +TIP body function remain speculative. One particular distinction is that Bik1 alone can phase separate, whereas Kar9 and Bim1 require additional factors to do so. This suggests that Bik1, particularly through its C-terminal EEY/F-like motif, plays a central role in determining the material properties of the +TIP body. Future studies will be needed to establish whether the fractal organization observed in Bik1 condensates is also present in the Kar9-Bim1-Bik1 complex in vivo. If this is the case, the high viscosity and mechanical resilience of the +TIP body may be critical for its ability to transmit force during mitosis and meiosis.
The methodology used in this study shows how the combination of SAXS and XL-MS allows to obtain molecular-level insights into protein phase separation. SAXS provides nanoscale information on protein conformation, while XL-MS identifies residue-specific interactions and conformational shifts under varying conditions. Together, these approaches offer a detailed view of both global and local structural changes, validating the detailed characterization of complex biomolecular condensates.
Ultimately, this study advances our understanding of the molecular mechanisms governing +TIP phase separation and provides a model for how proteins organize within condensed phases. As research into biomolecular condensation continues to expand, a fundamental question remains: How do cells harness phase separation to fine-tune dynamic cellular processes, and to what extent do aberrations in these systems contribute to human diseases?
Link to the Original Paper
Czub, M.P., Uliana, F., Grubić, T. et al. Phase separation of a microtubule plus-end tracking protein into a fluid fractal network. Nat Commun 16, 1165 (2025). https://doi.org/10.1038/s41467-025-56468-8
 (No Ratings Yet)
(No Ratings Yet)Get involved
Create an account or log in to post your story on FocalPlane.
More posts like this

Filter by
- NewsApply
- DiscussionsApply
- How toApply
- ToolsApply
- Case studiesApply
- InterviewsApply
- JobsApply
- EducationApply
- Blog seriesApply
- GloBIAS – updates fr..om the communityApply
- WAMBIAN: West Africa.. in FocusApply
- Volume EMApply
- Latin American Micro..scopistsApply
- Bio-image Analysis w..ith NapariApply
- Imaging with…Apply
- Towards Global Acces..sApply
- Latin America Bioima..gingApply
- From Zero to Qupath ..HeroApply
- Asian Microscopists ..and Cell BiologistsApply
- AIC at HHMI JaneliaApply
- Deep Learning for Bi..o-image analysisApply
- Highlights from Euro..-BioImagingApply
- LSFM seriesApply
- DIY MicroscopyApply
- View all