Compartmentalized Cytoplasmic Tradewinds
Posted by GalbraithLab, on 9 July 2026
Cathy Galbraith and Jim Galbraith share their story of answering how does a cell deliver a soluble protein — untethered to any motor, riding on no track — to the right place at the right time? They used six different imaging approaches to discover hidden cytoplasmic compartments and tradewinds that were hiding in plain sight.
Hidden in the Flow
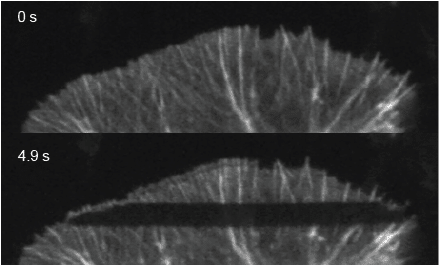
Because soluble proteins move so fast, they are hard to observe, and a default assumption has been that they move by diffusion: a rapid, random walk over short distances, with no sense of direction. Actin is one of the best examples of why this movement is so hard to see. Polymerized actin filaments treadmill backward from the leading edge. The monomers they shed must somehow get back to the front to be reused; otherwise, the cell would run out of raw material and stop protruding.
We know the movement occurs, but the journey is almost impossible to see. The retrograde flow of the structurally organized, optically dense polymerized actin network is the dominant, unmistakable motion. It moves roughly fifty times slower than the forward transport we were trying to measure, and its sheer presence dominates the visual field.
Ask any ordinary method “what’s moving here,” and it answers that the network is moving backward. That answer isn’t wrong. But it tells you nothing about the monomers heading the other way.
Six Methods, All Converging to the Same Answer
We designed a study with six independent approaches. Each one aimed at a different piece of the puzzle, so that no single instrumental quirk or labeling artifact could conceal an answer the others would independently uncover.
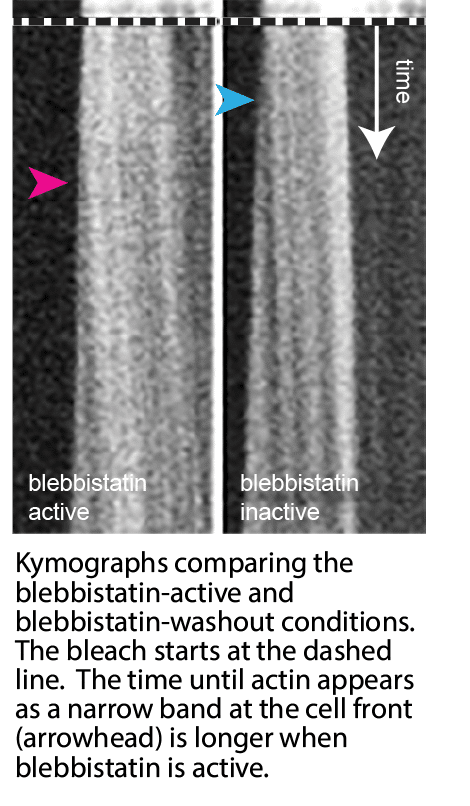
To start, bleach-labeling and kymography gave us a first raw velocity for the forward movement. This let us separate our hidden flow from the retrograde network treadmilling by exploiting their different timing. The forward velocity depended on myosin II. An optogenetic blebbistatin rescue let us test that directly: we silenced the motor with a drug, then switched its contraction back on by inactivating the drug with light, in the same cell. The fast forward transport reappeared.
Finding the Tradewinds
On its own, that result only told us that contraction was necessary. It didn’t tell us what kind of transport this was. The answer came from asking a different question: what else gets carried along?
Using our new photoactivation assay, FLOP (Fluorescence Leaving Original Point), and single-particle tracking, we found actin mutants that can’t polymerize move the same way. Even adhesion molecules and free dye that binds nothing at all move forward.
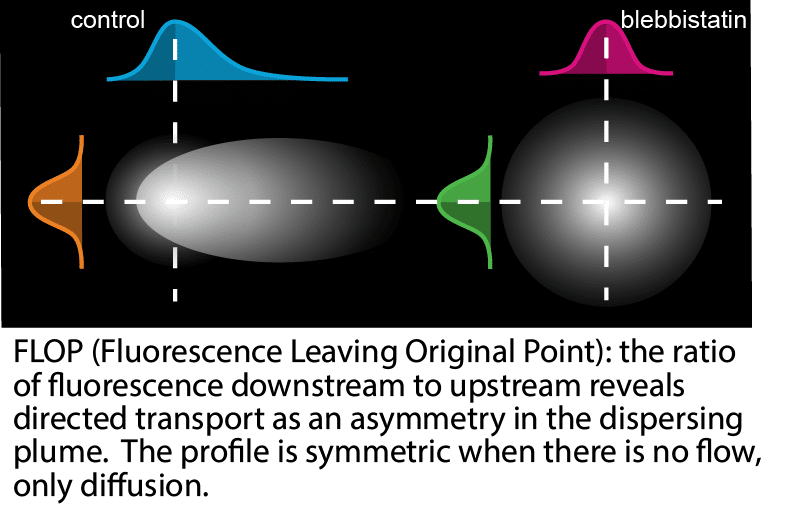
No motor drags all these proteins forward. And no track exists for them to ride on. Only one mechanism moves every one of those molecules, regardless of what they bind: bulk fluid flow. In other words, these are advection-driven cytoplasmic tradewinds. Fluorescence correlation spectroscopy, working from a completely different set of assumptions, yielded the same quantitative split between ordinary diffusion and the added advective flow.
Locating the Barrier
That led to another question: was this flow restricted to a defined region, or did it extend across the whole cell?
We turned to FLOP again, but instead of tracking the spread of a protein from a single point, we compared the spread of activated protein starting at three locations: inside the lamella, inside the cell body, or at the boundary between them. Protein starting in either compartment predominantly stayed where it started. Only protein starting right at the boundary spread into both compartments.
This demonstrated a discrete boundary, not just a flow that weakened with distance. We also found the barrier to be leaky rather than absolute, but FLOP couldn’t say what was doing the confining.
Seeing the Barrier Itself
For that, we turned to structured-illumination microscopy (SIM) and interferometric PhotoActivation and Localization Microscopy (iPALM).
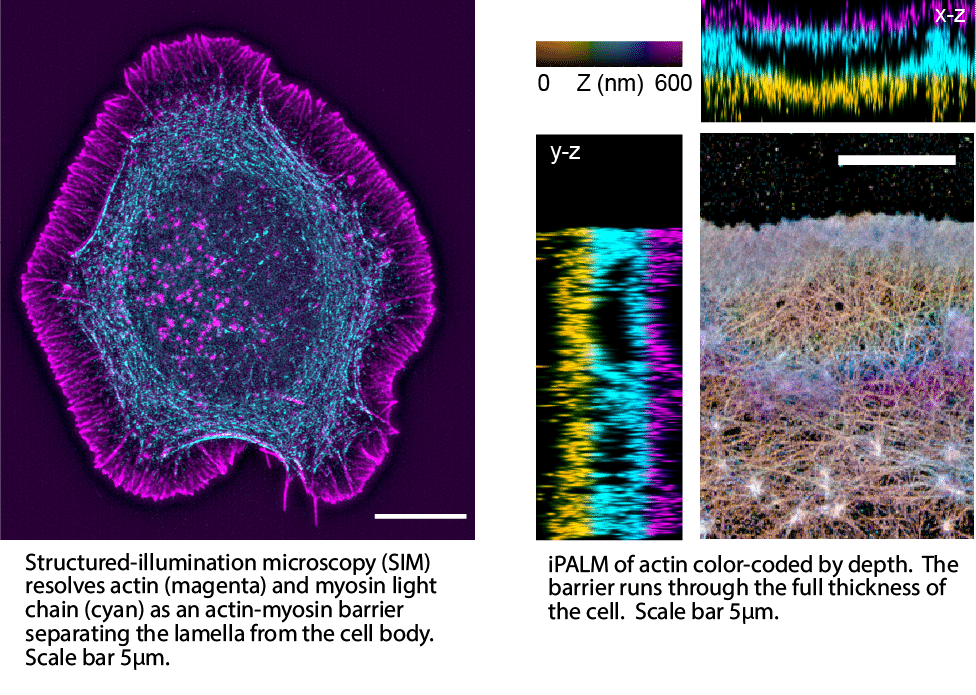
With the 15 nm resolution of iPALM, we found something nobody had seen before: a real, nanometer-resolved actin-myosin wall, sitting exactly where FLOP had located the boundary. In fact, it runs through the cell’s full thickness. This wall is the physical structure behind the leaky partition, and the source of the contraction driving fluid forward.
Six lines of evidence. Six blind spots overcome. One picture at the bottom of it all: a leaky actin-myosin barrier that partitions a compartment that generates its own internal cytoplasmic tradewinds.
The video below walks through that convergence, from the first look at retrograde flow to the barrier waiting at the end of it.
Why This Matters
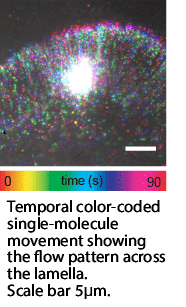
This compartment changes how we should think about the leading edge. It isn’t just the cytoskeleton, ratcheting up against the membrane. Instead, it’s a semi-enclosed region with its own internal flow. That flow concentrates and directs the exact proteins a protruding, adhering edge needs, moving them faster than diffusion ever could. It also steers that delivery toward whichever part of the edge is actively advancing.
A migrating cell, a cell closing a wound, an invading tumor cell — none of them are just polymerizing actin at the front. Rather, each one runs on a local, self-generated current that decides where the material actually goes.
We suspect this kind of directed transport, hidden underneath a larger and more easily imaged process, isn’t unique. Wherever it turns up next, it will matter for the same reason it matters here: cells that steer their own building blocks can respond faster and more precisely to their environment than cells relying on diffusion alone.
The Real Lesson
Getting there didn’t require a better microscope. It required stepping back, mapping the different biophysical layers of the phenomenon with a specific imaging technique, based on its unique ability to isolate each layer’s signal from the noise of the layer sitting on top of it. Anyone chasing their own small, hidden signal can apply that strategy directly, even where the underlying biology looks nothing like ours.
The full data, controls, and statistics behind every step above are in the paper, now out in Nature Communications.
Galbraith, C.G., English, B.P., Boehm, U., Galbraith, J.A. Compartmentalized cytoplasmic tradewinds direct soluble proteins. Nature Communications 17, 2589 (2026). https://doi.org/10.1038/s41467-026-70688-6
Acknowledgements
This work was supported by NIH grant GM117188 (CGG), NSF awards 171636 (JAG), 2345411 (CGG), W.M. Keck Foundation (CGG and JAG), and the Howard Hughes Medical Institute (BPE and UB). The iPALM work was partly supported by an award from the Advanced Imaging Center at Janelia. The SIM imaging was partly supported by a Core Research Facilities Grant from OHSU School of Medicine.
 (3 votes, average: 1.00 out of 1)
(3 votes, average: 1.00 out of 1)